
Regulation of Prokaryotic Gene Expression: Lactose operon
 المؤلف:
Denise R. Ferrier
المؤلف:
Denise R. Ferrier
 المصدر:
Lippincott Illustrated Reviews: Biochemistry
المصدر:
Lippincott Illustrated Reviews: Biochemistry
 الجزء والصفحة:
الجزء والصفحة:
 29-12-2021
29-12-2021
 2122
2122
Regulation of Prokaryotic Gene Expression: Lactose operon
The lac operon contains the genes that code for three proteins involved in the catabolism of the disaccharide lactose: the lacZ gene codes for β-galactosidase, which hydrolyzes lactose to galactose and glucose; the lacY gene codes for a permease, which facilitates the movement of lactose into the cell; and the lacA gene codes for thiogalactoside transacetylase, which acetylates lactose. [Note: The physiologic function of this acetylation is unknown.] All of these proteins are maximally produced only when lactose is available to the cell and glucose is not. [Note: Bacteria use glucose, if available, as a fuel in preference to any other sugar.] The regulatory portion of the operon is upstream of the three structural genes and consists of the promoter region where RNA pol binds and two additional sites, the operator (O) and the catabolite activator protein (CAP) sites, where regulatory proteins bind. The lacZ, lacY, and lacA genes are maximally expressed only when the O site is empty and the CAP site is bound by a complex of cyclic adenosine monophosphate ([cAMP] see p. 94) and the CAP, sometimes called the cAMP regulatory protein (CRP). A regulatory gene, the lacI gene, codes for the repressor protein (a trans-acting factor) that binds to the O site with high affinity. [Note: The lacI gene has its own promoter and is not part of the lac operon.]
1. When only glucose is available: In this case, the lac operon is repressed (turned off). Repression is mediated by the repressor protein binding via a helix-turn-helix motif (Fig. 2) to the O site, which is downstream of the promoter (see Fig. 33.4A). Binding of the repressor interferes with the binding of RNA pol to the promoter, thereby inhibiting transcription of the structural genes. This is an example of negative regulation.
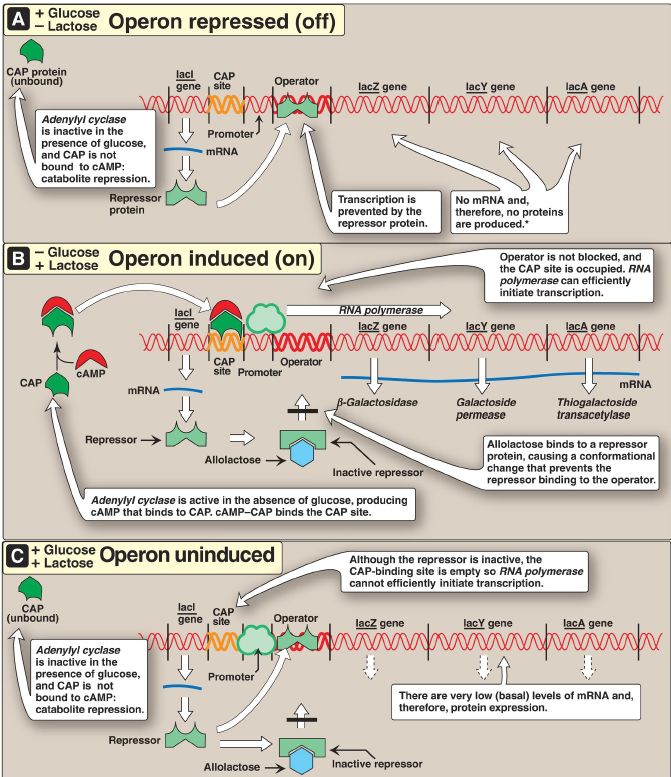
Figure 1: The lactose operon of Escherichia coli in the presence of A. only glucose, B. only lactose, and C. both sugars. *[Note: Even when the operon has been turned off, the repressor transiently dissociates from the operator at a slow rate, allowing a very low level of expression. The synthesis of a few molecules of permease (and β-galactosidase) allows the organism to respond rapidly should glucose become unavailable.] CAP = catabolite activator protein; cAMP = cyclic adenosine monophosphate; mRNA = messenger RNA.

Figure 2: Helix-turn-helix motif of the lac repressor protein.
2. When only lactose is available: In this case, the lac operon is induced (maximally expressed, or turned on). A small amount of lactose is converted to an isomer, allolactose. This compound is an inducer that binds to the repressor protein, changing its conformation so that it can no longer bind to the O site. In the absence of glucose, adenylyl cyclase is active, and cAMP is made and binds to the CAP. The cAMP–CAP transacting complex binds to the CAP site, causing RNA pol to initiate transcription with high efficiency at the promoter site (see Fig. 1B).
This is an example of positive regulation. The transcript is a single polycistronic mRNA molecule that contains three sets of start and stop codons. Translation of the mRNA produces the three proteins that allow lactose to be used for energy production by the cell. [Note: In contrast to the inducible lacZ, lacY, and lacA genes, whose expression is regulated, the lacI gene is constitutive. Its gene product, the repressor protein, is always made and is active unless the inducer is present.]
3. When both glucose and lactose are available: In this case, the lac operon is uninduced, and transcription is negligible, even if lactose is present at a high concentration. Adenylyl cyclase is inhibited in the presence of glucose (a process known as catabolite repression) so no cAMP–CAP complex forms, and the CAP site remains empty. Therefore, the RNA pol is unable to effectively initiate transcription, even though the repressor is not bound to the O site. Consequently, the three structural genes of the operon are expressed only at a very low (basal) level (see Fig. 1C). [Note: Induction causes a 50-fold enhancement over basal expression.]
 الاكثر قراءة في الكيمياء الحيوية
الاكثر قراءة في الكيمياء الحيوية
 اخر الاخبار
اخر الاخبار
اخبار العتبة العباسية المقدسة




















